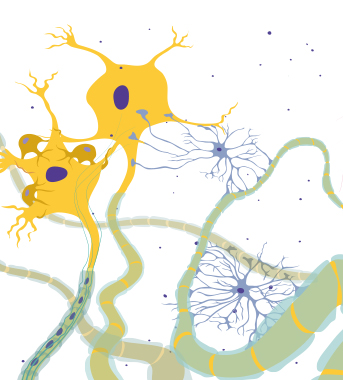
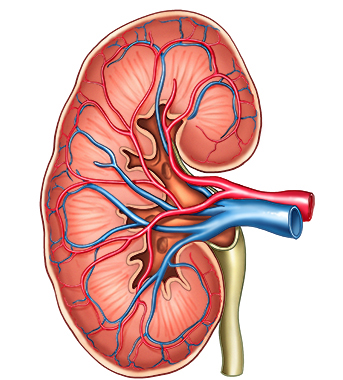
DIMEANE (English) Interactive Mobile E-learning Apps for European Nursing Education (DIMEANE) The DIMEANE project is part of the Erasmus+ program, and aims to develop interactive mobile e-learning applications for use in nursing education. Neurophysiology Nursing with focus on Cardiovascular diseases Nursing with focus on Temperature Regulation Anatomy and physiology of the heart Nursing with focus on skin and wound care Kidney anatomy and physiology Respiratory physiology Nutrition User guides